Atlas of Atlantic Ostracods
Taxon details
Halocypris inflata Dana, 1849
- Order:
- MYODOCOPA
- Suborder:
- Halocypridina
- Family:
- Halocyprididae
- Subfamily:
- Halocypridinae
- Size range (females):
- 1.4-1.8 mm
- Size range (males):
- 1.3-1.6 mm
- Depth:
- epi-/mesopelagic
Notes
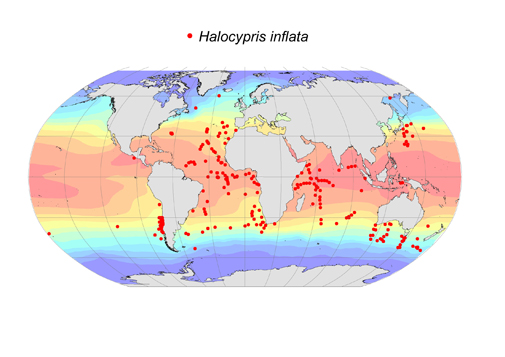
362 records
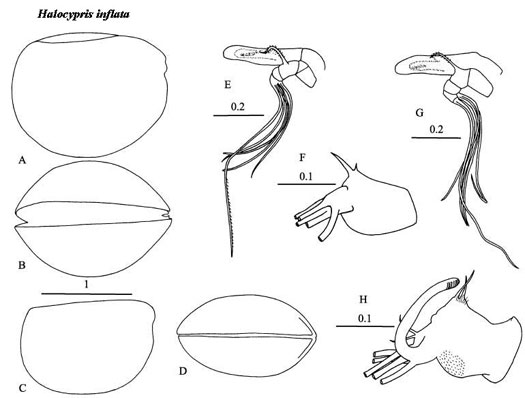
Recognizing members of this genus is straight forward because its globose species are unique in lacking a clearly developed rostrum. However, identifying them to species is of extreme difficulty, and this is reflected in the confusion associated with their systematics. Dana (1852) originally described two species H. inflata and H. brevirostris - probably male and female of the same species, but for neither ‘species’ was the description adequate. Claus (1874) named a new species H. concha, which was almost certainly synonymous with Dana’s ‘species’. Then Claus (1890) described a second species, H. pelagica distinguished on the basis of its smaller size. Müller (1906) synonymized the two size forms as H. inflata, but Skogsberg (1920) argued that the name H. brevirostris had precedence over H. inflata. Angel (1982) demonstrated that there are two species in the North Atlantic that co-occurred over limited sectors of their ranges. He argued the names H. inflata and H. pelagica were the most appropriate. Where the two species co-occur they are segregated bathymetrically, but not only did the larger inflata occupy slightly deeper water, but the carapace lengths of the juvenile instars became less variable. Angel (1982) suggested these responses may be an example of character displacement and hence be evidence of competition. Recently a new larger species has been recognized in the Pacific and is being described (Chavtur and Stovbun, in press).
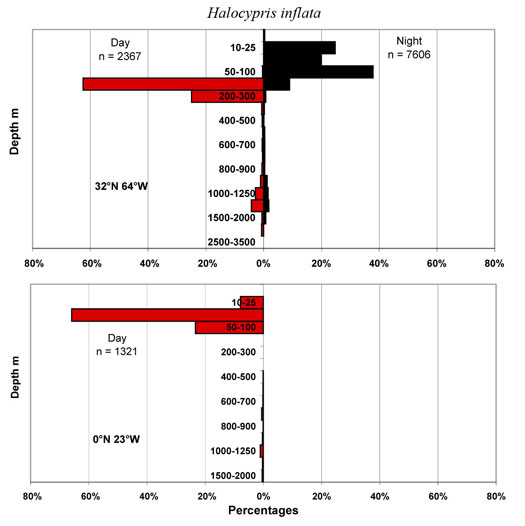
One result of all this confusion is that researchers have used either H. brevirostris or H. inflata without distinguishing the smaller species, H. pelagica. So the ranges for these two species shown in the maps may be totally unrepresentative. This, the larger of the two species, has been reported over a wide latitudinal range, 60°N to 54°S. When they have been distinguished the smaller species has a range limited to the tropics. There are a further 320 records of ‘brevirostris’ that cannot be ascribed to any of the presently recognized species. The species can be extremely abundant, sometimes dominating the epipelagic halocyprid populations. It undertakes diel vertical migrations at mid-latitudes and often occurs in abundance in the neuston at night.
| Equator | N | Mean mm | s.d. | Range mm |
|---|---|---|---|---|
| Female | 439 | 1.66 | 0.056 | 1.52-1.80 |
| Male | 389 | 1.45 | 0.043 | 1.32-1.60 |
| A-1 | 182 | 1.06 | 0.040 | 0.96-1.22 |
| A-2 | 114 | 0.74 | 0.022 | 0.70-0.80 |
| A-3 | 38 | 0.54 | 0.018 | 0.50-0.58 |
| Southern Ocean | N | Mean mm | s.d. | Range mm |
|---|---|---|---|---|
| Female | 138 | 1.49 | 0.076 | 1.22-1.70 |
| Male | ||||
| A-1 |
| Chavtur data
|
N | Mean mm | s.d. | Range mm |
|---|---|---|---|---|
| Female | 96 | 1.62 | 0.087 | 1.49-1.85 |
| Male | 163 | 1.45 | 0.055 | 1.33-1.55 |